解剖蛋白激酶脊柱以及如何打破它
磷酸基团向丝氨酸,苏氨酸和酪氨酸残基的翻译后添加是调节真核生物中蛋白质活性的基本策略。真核蛋白激酶 - 催化这些修饰的酶 - 对细胞功能至关重要,并且异常激酶活性与许多疾病相关,包括癌症,炎症,感染,糖尿病,高血压和神经变性。
因此,真核蛋白激酶是治疗干预的重要靶标,现在占所有药物发现和开发努力的四分之一,并且仅次于G蛋白偶联受体(GPCR)作为药物靶标。自2001年以来,已有超过三十种激酶抑制剂获得FDA批准,当时,bcr-abl激酶抑制剂伊马替尼(Gleevec)被批准用于治疗慢性粒细胞白血病(CML)。合理的药物设计在激酶抑制剂的开发中发挥了重要作用,分子被设计用于靶向特定的激酶构象。因此,了解调节蛋白激酶活性的结构基础是这些努力的关键。
对于PLOS Biology XV Collection,我选择突出Susan Taylor实验室的一篇文章(Meharena等,2013),该文章定义了一组分子间相互作用,区分了真核蛋白激酶的无活性和活性构象状态。这种分类是复杂的,因为与许多酶不同,真核激酶不具有单个离散的活性和非活性构象,而是动态的,具有填充两种功能状态的多种构象。
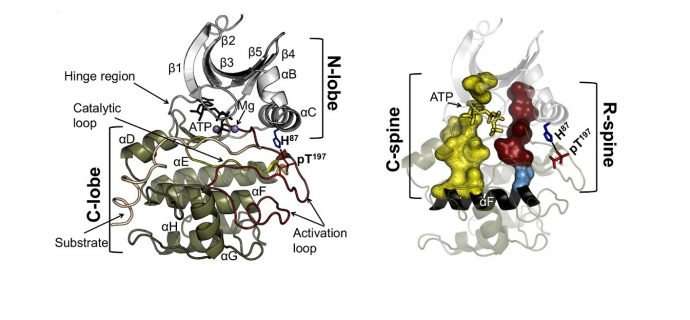
真核蛋白激酶的催化核心由保守的N-和C-叶组成,活性位点位于这两个叶的界面处。以前的研究已经确定了催化核心中的三个疏水特征:C-叶中的αF-螺旋和一级序列中的两个非连续残基簇,它们在三维结构中聚结形成两个疏水“刺”,跨越N-和C-叶。催化(C)脊柱包括结合ATP的腺嘌呤环,其桥接N-和C-叶中的疏水残基。Regulatory(R)脊柱,通常由C-叶中的两个芳香族残基(RS1和RS2)和N-叶中的两个脂肪族残基(RS3和RS4)组成,平行于C-脊柱,在处于活跃状态的连续疏水补片,
Meharena及其同事检测了约13,000种激酶序列中的R-脊柱残基,并通过对代表性激酶cAMP依赖性蛋白激酶(PKA)的突变分析测试了这些残基的性质假设。他们观察到C-叶中的残基RS1和RS2对突变非常敏感,这与N-叶中残基RS3和RS4的相对稳健性相反。这导致鉴定出围绕RS3和RS4的一组三个高度保守的残基。这些支持N-叶中R-脊柱的残基被称为壳残基Sh1,Sh2和Sh3。另外的突变分析提供了对R-脊柱完整性对催化必不可少的假设的实验验证。此外,
知道活性状态需要组装的R-脊柱,该小组检查了真核蛋白激酶的可用结构,并鉴定了其中拆解R-脊柱的172种结构。他们能够对R脊柱被破坏的四种特定方式进行分类。这些中的两个与先前表征的与活化环中DFG基序的定位相关的无活性构象相关。一种构象涉及DFG-out取向,另一种是由αC-螺旋的运动引起的DFG-in取向。这两种不活跃的构象已经成功地用于药物开发。
对其他非活性构象的描述为药物设计的新策略提供了机会,并为解释和最终调节由与人类疾病相关的蛋白激酶突变引起的分子缺陷提供了更广泛的基础。
推荐内容
-
不孕不育虫子的口水,却被当个宝贝卖?听听专家怎么说
随着互联网的快速发展,我们在获取信息的时候很容易被带跑方向。有时候真相也许并不是网上说的那样,所以我们必须要有自己的判断和独立...
-
17种“功效”全靠脑补,看螺旋藻“野鸡变凤凰”的故事!?其实是
互联网时代,每个人都是信息传播者,但是我们必须清楚一点:在没有获得真正证实的消息之前,千万不能随意散播谣言,很容易造成以讹传讹...
-
干细胞的发现获得诺贝尔医学奖
两名科学家发现,发育时钟可以在成熟细胞中逆转,将它们转化为不成熟细胞,使之具备成为体内任何组织的能力——多能干细胞。他们将获得...
-
得了甲状腺疾病一定要补碘吗?是真的吗?
互联网时代,每个人都是信息传播者,但是我们必须清楚一点:在没有获得真正证实的消息之前,千万不能随意散播谣言,很容易造成以讹传讹...
-
红壳鸡蛋营养价值更高吗?真是如此?
日常生活中我们经常会听到各种各样的坊间流言。然而很少人去求证这些流言的真实性,久而久之就变成了真事。在这里,小编必须提醒大家,...
-
这种激进的新方法用血管完整再生失败的肺
除了偶尔伴随跑步的灼痛外,大多数人都不会太注意在我们胸前喘气的双叶风琴。但肺部是工程奇迹的壮举:超过40种类型的细胞嵌入精致但柔...
-
提升老年人疫苗接种率仍具紧迫性
当前全国疫情防控处于攻坚阶段,基层防疫人员要加大力度反复宣传,说清楚老年人接种疫苗的重要性,消除老年人对可能引发不良反应的种种顾
-
发现血压基因
来自国际血压全基因组关联研究联盟的研究人员已经确定了总共23个与血压测量相关的基因区域。发表在NatureandNature Genetics上的两项研究
-
“会生气”的胰腺炎 夺命规律被揭示
科技日报讯 (记者俞慧友)4月25日,科技日报记者从中南大学湘雅医院获悉,历经多年临床观察和分析,该医院胰腺外科黄耿文教授团队在
-
吃蚕豆会得病?你知道吗
日常生活中我们经常会听到各种各样的坊间流言。然而很少人去求证这些流言的真实性,久而久之就变成了真事。在这里,小编必须提醒大家,...