消耗在海底发现的乙烷的难以捉摸的微生物
一股天然气从海底散发出来。这些碳氢化合物排放包含约90%的甲烷,其余大部分是乙烷。例如深海来源是负责甲烷,一种温室气体,存在于地球大气的5%1。通过捕获和使用这些气体作为其细胞碳和能量的唯一来源的微生物,从深海释放到大气中的烃的量显着减少。在自然中写作,陈等人。图2描述了为期十年的科学任务的最终结果,该任务旨在识别和表征造成深海碳氢化合物渗漏的乙烷消耗的微生物和反应。

已经确定了在没有环境氧气的情况下将甲烷,丙烷和丁烷的小烷烃分子代谢成二氧化碳的微生物(一种称为厌氧氧化的过程),但那些使用乙烷的微生物一直难以捉摸。尽管在环境氧(有氧氧化)存在下通过称为加氧酶的酶氧化烷烃是热力学上有利的反应,因此可能发生,但厌氧过程在热力学上是不利的。因此,只有当厌氧烷烃偶联到极热力学上有利的还原反应时才会发生厌氧烷烃氧化。
陈等人。表明乙烷的厌氧氧化涉及两种微生物之间的互利关系,称为“萎缩”(图1)。新鉴定的微生物,所述古细菌“暂定Argoarchaeum ethanivorans”,乙烷氧化成CO2。像所有已知的氧化甲烷,丙烷和丁烷的厌氧微生物一样。A.ethanivorans被归类为厌氧甲烷氧化(ANME)古细菌。陈等人。表明乙烷氧化成CO2需要另一种生化反应,即硫酸盐还原成硫化物。Ca的基因组。A. ethanivorans缺乏硫酸盐还原酶,但是对与Ca存在于相同微生物培养物中的微生物的分析。A.ethanivorans显示存在两种来自Desulfosarcina属的细菌菌株,其将硫酸盐还原成硫化物。
关于烷烃氧化古细菌和硫酸盐还原细菌之间的相互关系的性质存在激烈的争论。它是否涉及在伴侣物种之间交换代谢物,如氢分子,小有机分子或含硫分子?或者它是否涉及电子的直接种间转移?烷烃氧化微生物含有编码具有多个血红素基团的细胞色素酶的基因,支持电子直接转移的证据,催化还原和氧化反应3-5;这种反应涉及分子间的电子转移。此外,透射电子显微镜已经揭示了甲烷 - 或丁烷 - 氧化微生物和硫酸盐还原微生物之间存在丝状纳米线连接。这些长丝被认为是所需要的种间电子转移3,4。然而,对比证据还表明,参与海洋甲烷氧化的ANME古菌产生含硫分子,转移到硫酸盐还原伴侣微生物6。
陈等人。我认为,对于乙烷氧化,含硫代谢物的转移是最有可能构成这两种微生物之间相互作用的过程。这是因为Ca.A. ethanivorans在生长期间产生大量硫,缺乏在甲烷氧化微生物中观察到的纳米线结构,并且生长为单个细胞而不是生物膜(微生物细胞彼此粘附的结构)。所有这些特征都可以促进种间电子转移。但陈等人。还观察到他们研究的硫酸盐还原菌的基因组编码多血红素细胞色素和纳米线连接的组分类似于在一起生长的烷烃氧化和硫酸盐还原微生物的培养物中发现的那些3。因此,陪审团仍然不清楚这种种间耦合是如何发生的。
什么是乙烷转化成CO生化途径2?陈和他的同事毫不含糊地证明了Ca的基因组。A.ethanivorans包含三个编码先前未知酶的亚基的基因,我们称之为乙基辅酶M还原酶(ECR)。然后,他们使用质谱法鉴定ECR的蛋白质序列。ECR与称为甲基辅酶M还原酶(MCR)的酶密切相关,其存在于氧化或产生甲烷的微生物中。陈等人。还鉴定了由ECR,乙基辅酶M催化的反应的最终产物。
陈等人。模拟了Ca的3D结构。A. ethanivorans ECR和“的丁烷代谢酶(丁基-辅酶m还原酶)暂定Syntrophoarchaeum”4,并用的MCR的已知结构相比,它们Methanothermobacter marburgensis7。在这种结构比较的基础上,以及所有已知MCR相关蛋白序列的比对,他们得出结论,ECR和代谢非甲烷烃气体的其他酶在一组酶的总体内形成一个独特的簇。烷基辅酶M还原酶(ACR)家族,催化烷烃的厌氧氧化。
通过用MCR的充分研究的酶机制类似于8,它是可能的ECR通过转化成乙烷的乙基分子,这是非常具有反应性启动厌氧乙烷氧化。当研究人员能够产生足够量的ECR来确定其酶和生物物理特性时,这将是令人兴奋的。例如,当天然气在Ca处喷出时,它如何选择性地催化乙烷的氧化。A. ethanivorans生长的甲烷含量如此之高?作者的分析没有揭示ACR酶结合口袋的任何明显区别特征,包括ECR,这可以解释他们对代谢特定烷烃的偏好。然而,作者比较的结构代表ACR的非活动状态。希望这个酶家族的活性状态的晶体结构很快将被确定,以帮助澄清这个和其他有关其机制的问题。
陈等人。提出了从ECR产生的乙基辅酶M到CO2的合理途径。在该模型中,乙基辅酶M被转化为乙酰辅酶A,乙酰辅酶A是参与许多其他代谢过程的分子,然后通过称为反向Wood-Ljungdahl途径9的机制被氧化。作者使用基因组学和蛋白质组学方法鉴定了参与该途径的蛋白质。然而,乙基辅酶M向乙酰辅酶A的转化仍需要实验验证。对于其他非甲烷氧化途径也存在类似的知识差距,包括Ca的厌氧丁烷氧化。Syntrophoarchaeum4。
接下来的逻辑步骤是解决关于新鉴定的乙烷 - 氧化微生物与其硫酸盐还原配偶体之间的通讯性质的争议,并建立乙基辅酶M和乙酰辅酶A之间的代谢桥。
推荐内容
-
12月6日广东佛山疫情最新数据公布 广东昨日新增境外输入确诊病例1例
广东新增本土确诊1例。那么,对于广东疫情的这个话题,今天的你是否也在关注着?目前广东疫情什么情况?广东省新冠肺炎疫情情况12月5日0-24时
-
7月20日天津疫情最新数据公布 天津新增3例境外输入确诊病例
受新冠疫情影响,目前我国的疫情虽然得到了有效的控制,但国外的疫情依然十分严峻。7月18日18时至19日18时,天津无新增本土新冠肺炎确诊病
-
2019年《医学研究杂志》审稿专家名单(按姓氏拼音排列)
-2022年4月16日发(作者:阴虚体质的表现)2019年«医学研究杂志»审稿专家名单(按姓氏拼音排列)蔡 成 上海交通大学附属儿童医院曹 萍
-
99例分泌物检查结果临床分析
-2022年4月16日发(作者:湖南教育考试信息港)99例分泌物检查结果临床分析作者:赵晓红来源:《中外医疗》2011年第7期赵晓红上海市徐汇区虹
-
肩部装置销售力量Wright将击败并提高Q3
Wright Medical公布第三季度销售额增长14%,导致其超出分析师预期,并提升了今年的前景。杰弗里斯分析师表示,对强劲的季度很难找到错误,
-
B站起诉UP主获赔100万什么情况 B站为什么起诉UP主始末介绍
提起bilibili(简称B站)大家都不陌生了吧?创建于2009年6月26日,发展至今已有十年历史,这个以动漫、漫画、游戏内容为主的小破站可能曾经也
-
8月13日郑州疫情最新实时消息公布 郑州对机场和中高风险等区域人
机场,亦称飞机场、空港,较正式的名称是航空站。机场有不同的大小,除了跑道之外,机场通常还设有塔台、停机坪、航空客运站、维修厂等...
-
桂圆红枣鸡蛋糖水的功效作用
-2022年4月18日发(作者:健康证明怎么开)桂圆红枣鸡蛋糖水的功效作用桂圆红枣鸡蛋糖水是很常见的养生饮品之一,所以大多数的人都会想知道桂
-
肖战开启公益模式 用实际行动参与扶贫
起肖战,大家都不陌生。熟知肖战的人都知道,肖战其实之前是歌手出身,再加上本人颜值又高,而且多才多艺,能唱能跳!后来慢慢的转向演戏,
-
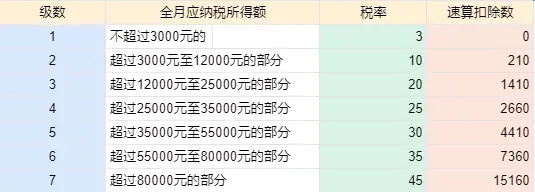
2022个人所得税税率表一览 2022年个人所得税计算方法是什么
个人所得税表是计算个人所得税的表。个人所得税率是个人所得税税额与应纳税所得额之间的比例。个人所得税率是由国家相应的法律法规规定...